Application Note FLEXstation3リーダーを用いた単一および
二波長インジケーターを用いた受容体を介した[Ca2+]iの変化のモニタリング
- 細胞内カルシウム、膜電位、細胞内pHの測定など、カイネティックな細胞ベース蛍光アッセイ用の汎用リーダー
- Gq共役型GPCRのモニタリングに使用されるカルシウム動員アッセイにおいて、信頼性が高く再現性の高いデータを得ることができる
- 色素のレシオメトリック特性により、シグナルは5倍に増加する
PDF版(英語)
はじめに
FlexStation® 3 マルチモードマイクロプレートリー ダーは、高感度光学系、流体移動、温度制御を 統合しており、細胞内カルシウム、膜電位、細胞内 pHの測定などのカイネティック細胞ベース フルオロメトリックアッセイに最適です。
カルシウム(Ca2+)は、バクテリアから特殊な神経細胞まで、細胞内で最も一般的なシグナル伝達要素です。受容体-リガンド相互作用のような様々な事象が、細胞内カルシウム([Ca2+] i )濃度をⒶ~100 nMからⒷ~1 µMに上昇させ、これがシナプス伝達物質の放出、筋収縮、ホルモン分泌、細胞運動、アポトーシスを含む多くの事象の引き金となる。
Ca2+] i )の変化は、Fluo-3やFLIPR® カルシウムアッセイキットなどの単波長色素またはFura-2などの二波長レシオメトリックインジケーターを用いて測定することができる。単波長色素では蛍光強度はCa2+濃度に依存し、レシオメトリック指示薬ではCa2+濃度に比例したスペクトルシフトが起こる(図1)。Fluo-3のような最も一般的な単一波長インジケーターは可視光で励起され、最も一般的な二波長色素(Fura-2)はUV光を必要とする2。
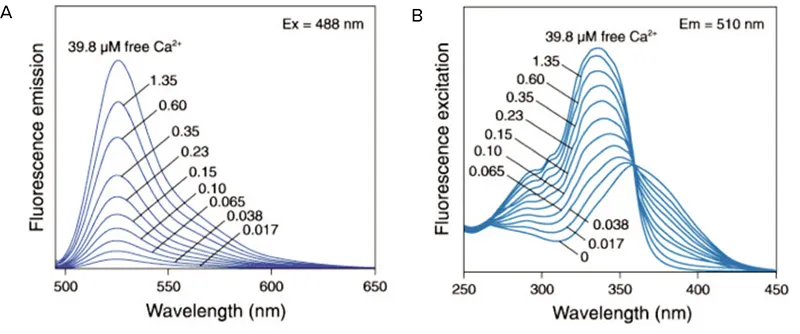
図1. Fluo-3の発光スペクトル。ゼロから39.8 µMの遊離Ca2+を封じ込めた溶液中でのFluo-3の蛍光発光スペクトル(A)とfura-2の蛍光励起スペクトル(B)。
** Molecular Probes Handbook of Fluorescent Probes and Research Chemicalsより転載。
FLEXstation 3リーダーには、調整可能なキセノンランプ ベースの光源とデュアル走査型モノクロメーターが組み込 まれており、機能的細胞アッセイに二波長色素を使用するこ とができます。可視光で励起可能なCa2+インジケーターは、サンプルの自動蛍光による干渉の低減や、色素による高い吸光度(より低濃度の色素の使用を可能にする)などの利点を提供するが、レシオメトリック色素は、従来の単一波長インジケーターよりもいくつかの利点を提供する。Fura-2の340/380 nm励起比を使用する利点:
- シグナルは色素濃度、照明強度、光路長に依存しない。
- 色素の不均一な装填、色素の漏れ、光漂白の影響を最小限に抑えることができる。通常、蛍光を著しく失うことなく、1時間にわたって検出を行うことができる。
- より安定したシグナルと、より優れたシグナル-ノイズ比が得られ、感度が向上する。
- 細胞内Ca 2+濃度の正確な測定が可能である。
別のアプローチとしては、FLIPRカルシウムアッセイキットなど、Molecular Devices社から入手可能な一連の「洗浄不要」試薬がある。これらもまた、従来のCa2+インジケーターに比べて多くの潜在的利点をもたらす。無洗浄」試薬は、従来のアッセイプロトコルにおけるこれらの問題を軽減する可能性がある:
- 洗浄手順中にプレーティングから除去されたセル
- 洗浄後の細胞の反応性(コンピテンス)の低下
- 洗浄バッファー残量のばらつき、リード化合物濃度のばらつき。
- 試験化合物の添加に伴う蛍光の低下(残存する細胞外色素の希釈による)。
このアプリケーションノートでは、FLEXstation 3 リーダーで一波長および二波長の Ca2+インジケーターを使用することと、ホモジニアスアッセイ システムの可能な利点を示します。
背景
血小板の調製
CHO-K1細胞には内因性ヌクレオチド受容体(P2Y2)3,4が存在し、ホスホリパーゼCの活性化、イノシトールリン酸(IP)の生成、プロテインキナーゼC(PKC)の活性化、細胞内Ca2+貯蔵の放出に関与しています。P2X受容体はATPゲートイオンチャネルであるのに対し、P2Y受容体はGPCRのスーパーファミリーに属します。P2Y受容体ファミリーには、P2Y1、P2Y2、P2Y4、P2Y6、P2Y11、P2Y12、P2Y13、P2Y14受容体が含まれます6。P2Yサブタイプは、アデニンとウラシルのヌクレオチドに対する選択性において薬理学的に異なっており、P2Y2受容体に対する主な生理学的リガンドはUTPとATPです7。P2Y2-受容体の重要な機能的役割の一つは、気道上皮における塩化物イオンのフラックスと肺の粘膜繊毛クリアランスの制御であり、嚢胞性線維症やその他の気道障害に関与している可能性がある。
CHO-K1細胞には内因性ヌクレオチド受容体(P2Y2)3,4が存在し、ホスホリパーゼCの活性化、イノシトールリン酸(IP)の生成、プロテインキナーゼC(PKC)の活性化、細胞内Ca2+貯蔵の放出に関与しています。P2X受容体はATPゲートイオンチャネルであるのに対し、P2Y受容体はGPCRのスーパーファミリーに属します。P2Y受容体ファミリーには、P2Y1、P2Y2、P2Y4、P2Y6、P2Y11、P2Y12、P2Y13、P2Y14受容体が含まれます6。P2Yサブタイプは、アデニンとウラシルのヌクレオチドに対する選択性において薬理学的に異なっており、P2Y2受容体に対する主な生理学的リガンドはUTPとATPです7。P2Y2-受容体の重要な機能的役割の一つは、気道上皮における塩化物イオンのフラックスと肺の粘膜繊毛クリアランスの制御であり、嚢胞性線維症やその他の気道障害に関与している可能性がある。
CHO-K1細胞には内因性ヌクレオチド受容体(P2Y2)3,4が存在し、ホスホリパーゼCの活性化、イノシトールリン酸(IP)の生成、プロテインキナーゼC(PKC)の活性化、細胞内Ca2+貯蔵の放出に関与しています。P2X受容体はATPゲートイオンチャネルであるのに対し、P2Y受容体はGPCRのスーパーファミリーに属します。P2Y受容体ファミリーには、P2Y1、P2Y2、P2Y4、P2Y6、P2Y11、P2Y12、P2Y13、P2Y14受容体が含まれます6。P2Yサブタイプは、アデニンとウラシルのヌクレオチドに対する選択性において薬理学的に異なっており、P2Y2受容体に対する主な生理学的リガンドはUTPとATPです7。P2Y2-受容体の重要な機能的役割の一つは、気道上皮における塩化物イオンのフラックスと肺の粘膜繊毛クリアランスの制御であり、嚢胞性線維症やその他の気道障害に関与している可能性がある。
Z'因子分析8とEC50推定値およびシグナルの大きさの比較を用いて、従来のCa2+インジケーターよりもレシオメトリックCa2+色素およびホモジニアスアッセイフォーマットの利点を示す。
材料
- セル CHO細胞-接着細胞(ECACC, Cat.#85051005)
- 試薬 FLIPR Calcium Assay Kit(Molecular Devices、Cat. #R8041)、ATP(アデノシン5'-三リン酸二ナトリウム塩、Sigma、Cat. #A2383)、プロベネシド(Sigma、Cat. #P8761)、DMSO、低含水(Sigma、Cat. #D2650)、Fluo-3 AMエステル(Molecular Probes、Cat. #F1242)、Fura-2 AMエステル(Molecular Probes、Cat. #F1221)、20%プルロン酸溶液(Molecular Probes、Cat. #P3000)
- プラスチック器具:75 cm 2培養フラスコ(Corning、 Cat. #430641)、黒壁透明底96ウェルプレート(Costar、 Cat. #3603)、透明U底96ウェルポリプロピレンプレート (Greiner Bio-One、Cat. #650101)、FlexStation用ピペットチップ - 200 μL非滅菌ポリプロピレンチップ(Molecular Devices、黒色Cat. #9000-0911または透明Cat. #9000-0912)
- 培地とバッファー Hams-F12 培地(Invitrogen Life Technologies 社、Cat. #31765-027)、Hank's Balanced Salt Solution (10X)(Invitrogen Life Technologies 社、Cat. #14065-049)、HEPES 1M 緩衝液(Invitrogen Life Technologies 社、Cat. #15630-056)、FBS:(Fetal Bovine Serum, heat inactivated, Invitrogen Life Technologies 社、Cat. #10099-141)、Geneticin(G418、Invitrogen Life Technologies、Cat. #10131-019)
方法
Fluo-3およびFura-2試薬の調製
洗浄バッファー:このバッファーには、滅菌したd.H2Oで希釈した1X HBSSと20mM HEPESの最終濃度を封じ込めた。重要:陰イオン交換タンパク質による色素の押し出しを防ぐため、ローディング培地には2.5mMのプロベネシドを添加した。
250mMのプロベネシドの新しいストックを調製する。710 mg のプロベネシドを 5 mL の 1 M NaOH に溶かし、5 mL の洗浄バッファー を加えて激しく撹拌する。
Fluo-3 と Fura-2 のストック溶液。50 µg の Fluo-3 AM または Fura-2 AM に 25 µL の DMSO を加える(最終濃度 Ⓐ~2 mM)。
色素ローディングバッファー 25μLのFluo-3またはFura-2と25μLの20%プルロン酸溶液を混合し、これを2.5mMのプロベネシドを封じ込めた12mLの洗浄液に加え、120μLのFBSを加える。これで96ウェルプレート約1枚分の試薬が調製できる。プルロニック酸は色素の溶解性と取り込みを促進するために使用されます。
FLIPR カルシウムアッセイキットの調製
キット添付の説明書に記載されているように、コンポーネント B を用いて 1X 試薬バッファー を調製し、コンポーネント A を 1X 試薬バッファーで希釈してローディングバッファーを調製する。
上記のように 250 mM Probenecid の新しいストックを調製する。
250 mM Probenecid の 1:50 希釈液を Loading Buffer(最終濃度 5 mM)に、250 mM Probenecid の 1:100 希釈液を 1X Reagent Buffer(最終濃度 2.5mM)に添加する。
セル調製
CHO細胞(継代番号<40)を、10%ウシ胎児血清と5 µg/mL G418を添加したHams-F12培地中で、75cm2のフラスコで細胞単層として培養し、CO2インキュベーター内で37℃に維持した。セルは2-3日ごとに1:10で継代した。
Ca2+] iの測定
CHO細胞を、上記のように添加したHams-F12培地中で、黒壁のクリアベース96ウェルプレート(96ウェルCostar社製)に1ウェル当たり3万個の密度で播種し、一晩培養した。その後、細胞を適切なカルシウム指示薬(Fluo-3、Fura-2、FLIPR Calcium Assay Kit)、および2.5 mMプロベネシドとともに37℃で60分間インキュベートした(付録参照)。
Fluo-3またはFura-2を負荷したセルは、2.5 mMプロベネシドを封じ込め たハンクスBSS培地で3回手動で洗浄した(BioHit 8-channelピペット)。各ウェルの最終容量は200 µLであった。
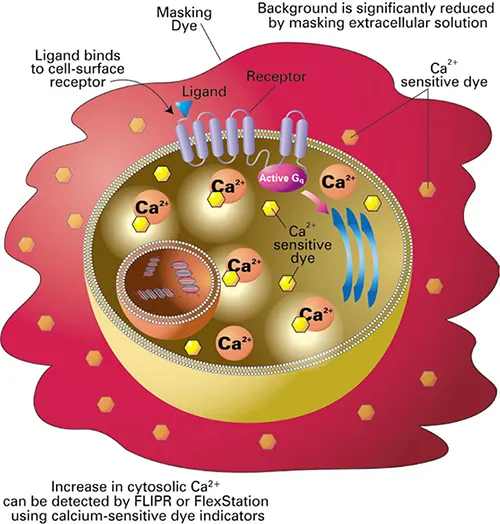
図2. FLIPRカルシウムアッセイの原理。FLIPRカルシウムアッセイキットにはカルシウム感受性色素が含まれており、インキュベーション期間中に細胞質内に取り込まれる。細胞不透過性のマスキング色素は細胞外に留まり、バックグラウンドの蛍光を遮断する。リガンドがレセプターに結合すると、カルシウムが細胞質内に放出される。色素は細胞内カルシウムに結合し、蛍光の増加が測定される。
次にセルプレートをFlexStation 3リーダー にセットし、ATP添加前後の蛍光をモニターした(50µL 添加、ATPは最終濃度5X、最終濃度30 nm- 30 µM)。実験パラメーターを以下に示す:
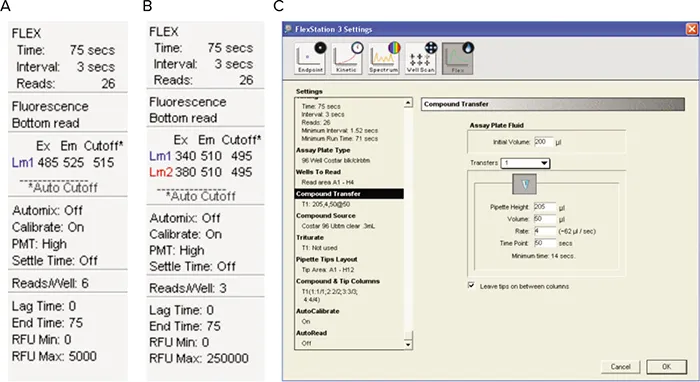
図3. SoftMax® Proソフトウェアの設定。(A)単一波長色素のみ、(B)Fura-2のみ、(C)All 3色素の実験パラメータ。
データ解析
反応は、ピーク蛍光強度から基底蛍光強度を引いた値(Fluo-3 または FLIPR Calcium Assay Kit)、または Fura-2 の場合は SoftMax Pro 内で計算した 340/380 nm 励起比として測定した。データはn > 12の平均±s.e.平均で表した。
E/[A]カーブデータの個々のセットは、4パラメータのロジスティックにフィット処理した:
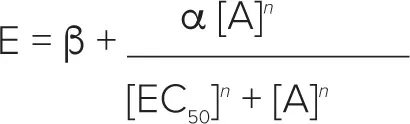
ここで、a、b、EC50 および n はそれぞれ、漸近線、曲線の底、位置(-log10 EC50 として表現)および傾きのパラメーターである。すべてのカーブフィッティング・ルーチンは、GraphPad Prism 3.0(GraphPad Software Inc.)
Z´因子は次式で計算した:
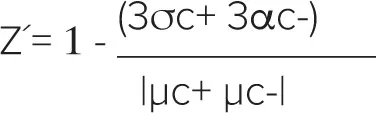
ここでσc+は陽性コントロールの標準偏差を表し、σc-は陰性コントロールの標準偏差を表す。μc+-μc-|は陽性コントロールの平均値と陰性コントロールの平均値の差の絶対値を表す。Z´ factor > 0.5は陰性コントロールと陽性コントロールの間に大きな分離帯があり、優れたアッセイであることを示す。
結果
ATPは、Fluo-3、Fura-2およびFLIPRカルシウムアッセイキットをそれぞれ負荷した細胞で、同様のEC50値を示しながら、CHO細胞において濃度依存的に[Ca2+]iの増加を惹起した(図4.
ATPは、Fluo-3、Fura-2およびFLIPRカルシウムアッセイキットをそれぞれ負荷した細胞で、同様のEC50値を示しながら、CHO細胞において濃度依存的に[Ca2+]iの増加を惹起した(図4.
| Fluo-3 AM | Fura-2 AM | FLIPR 試薬 | |
|---|---|---|---|
|
EC 50 |
6.39 ± 0.12 | 6.44 ± 0.13 | 6.51 ± 0.07 |
| シグナル(倍) | 1.8 X | 4.8 X | 5.6 X |
| Z'因子 | 0.28 | 0.60 | 0.68 |
表1. CHO細胞におけるATP E/[A]曲線から算出した3つのパラメーターの比較: EC50、蛍光シグナルの増加倍率、およびZ'ファクター。
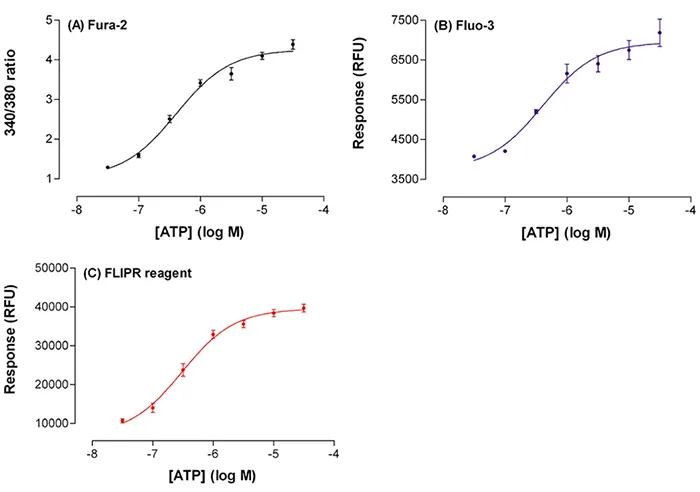
図4. 蛍光シグナルによるATPの増加。Fluo-3(B)とFLIPRカルシウムアッセイキット(C)、またはFura-2の340/380 nm励起比(A)を負荷した細胞の蛍光シグナルの増加(y軸)に対するATP濃度(x軸)のプロット。Fura-2比は、SoftMax® Proソフトウェアで時間補間したデータから算出した。データは平均±s.e.平均。
これらの結果は、3種類の蛍光試薬を負荷したセルからのATP EC50値に無視できるほどの差がないことを示している。しかし、2つの単一波長試薬の蛍光強度のピークには有意な差があった。Fluo-3では、シグナルが基底値より1.8倍増の3,200RFUになったのに対し、FLIPRカルシウムアッセイキットでは5.6倍増の32,000RFUになった。ピーク反応はFluo-3ではより変動しやすい傾向があった(図4参照)。Fura-2では、シグナルは基底値より4.8倍増加した。
シグナルのダイナミックレンジとアッセイのデータ変動を反映するスクリーニング係数ウィンドウ(Z'ファクター)8は、バッファー添加(ネガティブコントロール)と 30 µM ATP 添加データ(ポジティブコントロール)を用いて計算した。Fura-2とFLIPRカルシウムアッセイキットで得られたZ´因子はそれぞれ0.60と0.68であった(表1)。これは陰性対照と陽性対照の間に大きな分離帯があり、再現性の高い高品質なアッセイであることを示している。Fluo-3のデータで得られたZ'ファクターは0.28で、「使用可能な」アッセイであったが、これはおそらくアッセイのプロトコールが最適でなかったことと、その後のアッセイのダイナミックレンジが小さかったことを反映していると思われる。
結論
本研究では、P2Y2-受容体を内因的に発現するCHOセ ルにおいて、ATPは以前に報告されたものと同様の効力 をもって、濃度依存的に[Ca2+] iの増加を引き起こした。さらに、FLEXstation 3リーダーを用いることで、このようなGq-共役GPCRをモニターするためのカルシウム動員アッセイにおいて、信頼性が高く再現性のあるデータを作成できることを実証した。
ATPのEC50推定値は、蛍光色素Fluo-3(6.39)、Fura-2(6.44)、FLIPRカルシウムアッセイキット(6.51)のいずれを用いても同程度であり、同じ日のアッセイプレート間でも、異なる日に分析したアッセイ間でも一貫していた(0.6 log unit以下の差)。しかし、3つの異なるローディングプロトコールによるダイナミックレンジには有意差があり、これは対応するZ'因子に反映されている。細胞をFluo-3で負荷した場合、シグナルは基底値の2倍以下の増加しか観察されなかったが、その最も可能性の高い原因は、プロトコルのその後の洗浄工程が細胞の反応性を低下させたことである。より低い継代番号のセルを使い、専用の細胞洗浄器(AquaMax® Washerまたは同様のもの)を使えば、おそらくFluo-3を負荷した細胞のZ'ファクターは改善されたであろう。これらの実験条件下で、Fura-2では、細胞が何度も洗浄ステップを経たにもかかわらず、シグナルが5倍増加した。これは、この色素の有益なレシオメトリック特性によるものと考えられる。FLIPRカルシウムアッセイキットでシグナル、ひいてはZ'ファクターの最も大きな倍率の増加が観察されたことから、この「無洗浄」ホモジニアスアッセイが、より強いシグナル、より高いアッセイ精度、生理学的データの改善につながることが示された。
FLEXstation 3 リーダーは、薬理学や薬剤スクリーニングのラボに使いやすく多用途のツールを提供し、細胞内カルシウム、膜電位、細胞内pHの測定のようなカイネティック、細胞ベースのフルオロメトリックアッセイに適していることが証明されています。
Molecular Devices社のホモジニアスアッセイキットは、アッセイにお けるより少ないステップで同じ目標を達成することが証明さ れていますが、FLEXstation 3 リーダーはレシオメトリック染料を使 用できるため、データの忠実度の向上に役立ちます。
参考文献
- Berridge, M.J. (1997). カルシウムシグナルの初歩的側面とグローバルな側面。J. Physiol., 499, 291 - 306.
- Haughland, R.P. (2000). 分子プローブ蛍光プローブと研究用化学物質のハンドブック。第20章 Ca 2+ , Mg 2+ , Zn 2+ およびその他の金属イオンの指標。
- Agi Schonbrunn, Ph.D., Department of Integrative Biology, Pharmacology and Physiology, University of Texas Health Science Center, Houston, Texasによる内因性GPCRリスト。(https://www.tumor-gene.org/GPCR/gpcr.html)
- Strassheim, D. and Williams, C.L. (2000). P2Y2 Purinergic and M3 Muscarinic Acetylcholine Receptors Activate Different Phospholipase C- Isoforms Are Uniquely Susceptible to Protein Kinase C-dependent Phosphorylation and Inactivation. J. Biol. Chem., 275, 39767- 39772.
- Conant, A.R. et al (1998). ヒト臍帯静脈内皮細胞株ECV304上のP2受容体の特性化。Br. J. Pharmacol., 125, 357-364.
- Geoffrey Burnstock, Maria-Pia Abbracchio, Jean-Marie Boeynaems, José L. Boyer, Stefania Ceruti, Marta Fumagalli, Christian Gachet, Rebecca Hills, Robert G. Humphries, Kenneth A. Jacobson, Charles Kennedy, Brian F. King, Davide Lecca, Maria Teresa Miras-Portugal, Gary A. Weisman. (2010). P2Y受容体。IUPHAR データベース (https://www.iuphar-db.org/ DATABASE/FamilyMenuForward?familyId=52).
- Law, C., Lydford, S.J. & Henderson, D.J. (1998). FLIPRを用いたECV304内皮細胞上のP2受容体の特性解析。NaunynSchmeidebergs Arch. Pharmacol. 358, R126.
- Zhang, J-H., Thomas, T.D.Y. and Oldenburg, K. R. (1999). ハイループットスクリーニングアッセイの評価とバリデーションに使用するシンプルな統計パラメータ。J. Biomol. Screening, 4, 67-73.
FlexStation 3 マルチモードマイクロプレートリーダーについてさらに詳しく>>
PDF版(英語)